

 Курсовая работа: Пептиды и первичная структура белка
Курсовая работа: Пептиды и первичная структура белка
Курсовая работа: Пептиды и первичная структура белка
ПЕПТИДЫ И ПЕРВИЧНАЯ СТРУКТУРА БЕЛКА
СОДЕРЖАНИЕ
ВВЕДЕНИЕ
ЛИТЕРАТУРНЫЙ ОБЗОР
1.1 Понятие о пептидах. Их строение
1.2 Номенклатура пептидов
1.3 Основной принцип пептидного синтеза
1.4 Экспериментальные методы создания пептидной связи
1.5 Первичная структура белка
1.6 Экспериментальное определение первичной структуры белка
ВЫВОДЫ
СПИСОК ИСПОЛЬЗОВАННОЙ ЛИТЕРАТУРЫ
ВВЕДЕНИЕ
Пептиды и белки представляют собой высокомолекулярные органические соединения, построенные из остатков α-аминокислот, соединенных между собой пептидными связями.
Ни один из известных нам живых организмов не обходится без белков. Белки служат питательными веществами, они регулируют обмен веществ, исполняя роль ферментов – катализаторов обмена веществ, способствуют переносу кислорода по всему организму и его поглощению, играют важную роль в функционировании нервной системы, являются механической основой мышечного сокращения, участвуют в передаче генетической информации и т.д. Как видно, функции белков в природе универсальны. Белки входят в состав мозга, внутренних органов, костей, кожи, волосяного покрова и т.д. Основным источником α - аминокислот для живого организма служат пищевые белки, которые в результате ферментативного гидролиза в желудочно-кишечном тракте дают α - аминокислоты. Пептиды и белки различают в зависимости от величины молекулярной массы. Условно считают, что пептиды содержат в молекуле до 100 (соответствует молекулярной массе до 10000), а белки - свыше 100 аминокислотных остатков (молекулярная масса от 10000 до нескольких миллионов). При этом в пептидах различают олигопептиды, содержащие в цепи не более 10 аминокислотных остатков, и полипептиды, содержащие до 100 аминокислотных остатков.
ЛИТЕРАТУРНЫЙ ОБЗОР
1.1 Понятие о пептидах. Их строение
Пептиды — это цепочечные молекулы, содержащие от двух до ста остатков аминокислот, соединенных между собой амидными (пептидными) связями.
|
|
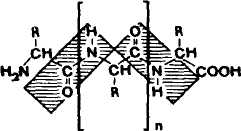
Рис.1 Строение пептида
Термин «пептиды» был предложен известным химиком Эмилем Фишером (1852—1919 гг.). Слово образовано из первых четырех букв названия пептоны (продукты расщепления белков пепсином) и конечных букв названия углеводов полисахариды.
По размеру молекулы и своим свойствам пептиды стоят между высокомолекулярными белками и аминокислотами. Наиболее распространены линейные пептиды, однако известны также циклические пептиды, молекулы которых могут иметь различные размеры. Циклические пептиды образуются из линейных, когда пептидная связь связывает амино- и карбоксильную функцию N- и С-концевых аминокислот.

Полинг и Кори в 1951 г. показали с помощью рентгеноструктурного анализа аминокислот, амидов аминокислот и простых линейных пептидов, что пептидная связь С—N укорочена по сравнению с нормальной простой связью (рис. 2).
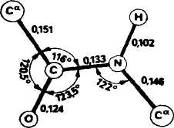
Рис. 2. Средние расстояния между атомами (нм), образующими пептидную связь и углы между связями.
Вследствие мезомерии получаются две устойчивые плоские конформации, транс (1) и цис (2), при затрудненном свободном вращении около связи С—N:
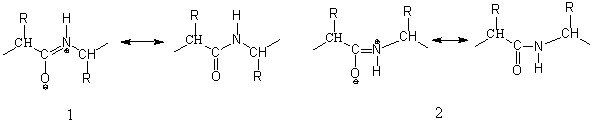
Рис.3 транс (1) и цис (2) пептиды
В 2,5-дикетопиперазинах, простейших циклических пептидах, построенных из двух аминокислот, имеются цис-пептидные связи. Циклические трипептиды могут существовать без напряжения также только с тремя цис-пептидными связями. Поскольку пролин и саркозин не обладают возможностью стабилизации транс-пептидиой связи, то можно легко синтезировать циклический трипептид — циклотрипролил. В нативных пептидах и белках преобладает транс-пептидная связь. В некоторых белках были найдены также и цис-пептидные связи, при этом в образовании пептидной связи всегда участвовал пролин [1].
1.2 Номенклатура пептидов
По числу аминокислот, содержащихся в пептиде, различают ди-, три-, тетра-, пента-, ..., окта-, нона-, декапептиды и т. д. Чтобы избежать проблемы, связанной с греческой нумерацией длинноцепочечных пептидов, Бодански предложил количество аминокислотных остатков пептида обозначать арабской цифрой и помещать перед словом «пептид». Например, 7-пептид вместо гептапептид, 10-пептид вместо декапептид. Пептиды, в молекулах которых меньше десяти аминокислотных остатков, формально относятся к олигопептидам, пептиды, построенные из большего числа аминокислотных остатков (до — 100),— к полипептидам. Различие между полипептидами и белками (макропептидами) чрезвычайно проблематично. Исторически сложилось так, что границей между полипептидами и белками считают соединения с молекулярной массой ~ 10 000, т. е. состоящие примерно из 100 остатков аминокислот. Такой принцип классификации основан на способности к диализу через природные мембраны[2].
Согласно принципам рациональной номенклатуры, пептиды рассматривают формально как ациламинокислоты, причем аминокислоте, карбоксил которой участвует в пептидной связи, придается окончание-ил. Поэтому только С-конечная аминокислота сохраняет свое первоначальное тривиальное название. По предложению Бейли, в формулах линейных пептидов аминокислота со свободной аминогруппой называется N-концевой аминокислотой, в горизонтально изображенной пептидной цепи она стоит слева. Аминокислота со свободной карбоксильной группой обозначается как С-концевая аминокислота. Фромажо предложил остаток, несущий сободную α-аминогруппу, называть начальной аминокислотой, а соответствующий остаток со свободной карбоксильной группой — конечной аминокислотой. Хотя это предложение кажется более простым, широкое признание получила рекомендация Бейли.
Число и последовательность связанных в пептид аминокислот называют первичной структурой. Для пептида с известной последовательностью формулу записывают, соединяя символы аминокислотных остатков черточками. Наконец, различают собственно пептид, например Ala-Ser-Asp-Phe--Gly и фрагмент -Ala-Ser-Asp-Phe-Gly- (с черточками при концевых аминокислотах). Если часть последовательности пептида еще не известна, то аббревиатуры соответствующих аминокислот, разделенные (запятыми) указывают в скобках:
Gly-Gly-Ala-Ser-Phe-(Tyr, Phe, Pro, Arg, Lys)-Val-Pro-Gly-Ala
1.3 Основной принцип пептидного синтеза
Образование пептидной связи в случае дипептида является простым химическим процессом. Дипептид формально получается при отщеплении молекулы воды от амино- и карбоксильной групп двух аминокислот (рис. 4). Последовательное повторение этого процесса, казалось бы, должно привести к длинным пептидам и даже к белкам. Однако реализация этого принципа возможна только в жестких условиях неконтролируемой реакции. Основатель пептидной и белковой химии Э. Фишер в 1906 г. писал: «Если бы сегодня по счастливой случайности с помощью какой-то жесткой реакции, например при сплавлении аминокислот в присутствии водоотнимающих средств, удалось получить настоящий белок и если бы, что еще менее вероятно, можно было искусственно созданный продукт идентифицировать с естественным, то это ничего не дало бы ни для химии белков, ни для биологии».

Рис.4 Основной принцип пептидного синтеза
Образование пептидной связи в мягких условиях удается лишь при активировании карбоксильного компонента одной из аминокислот, вступающей в реакцию (рис. 5).
Вторая аминокислота В (аминокомпонент) атакует активированный карбоксильный компонент аминогруппой с образованием пептидной связи. Незащищенная аминофункция карбоксильного компонента А тоже может реагировать, что приводит (рис. 5) к нежелательным побочным продуктам — линейным и циклическим пептидам. Из этого следует вывод, что для однозначного течения пептидного синтеза следует временно блокировать все функциональные группы, не участвующие в образовании пептидной связи[3].

Рис.5 Схема образования пептидной связи без защиты не участвующих в реакции функциональных групп
Пептидный синтез, т. е. образование каждой пептидной связи, является поэтому многоступенчатым процессом. В первую очередь получают частично замещенные аминокислоты, при этом они одновременно теряют цвиттер-ионную структуру. Вторая ступень, собственно образование пептидной связи, протекает в две стадии. Сначала нужно активировать N-защищенный карбоксильный компонент. Затем происходит собственно образование пептидной связи, которое протекает либо одноступенчато (вместе с активированием), либо последовательно в следующую стадию. На третьей ступени защитные группы селективно отщепляются, причем полученные частично защищенные производные дипептидов могут использоваться для дальнейших синтезов как карбоксильные или аминокомпоненты. Само собой разумеется, что в случае синтеза дипептида обе защитные группы удаляются одновременно. Пептидный синтез, далее, усложняется еще и тем, что из 20 протеиногенных аминокислот 9 обладают еще третьей функциональной группой, которая также требует селективной защиты. Это Ser, Thr, Туг, Asp, Glu, Lys, Arg, His и Cys. Следует различать временные и постоянные защитные группы. Временные защитные группы служат для защиты концевых амино- и карбоксильных групп и должны поэтому селективно отщепляться в присутствии постоянных защитных групп. Постоянные защитные группы удаляются обычно только после окончания синтеза пептида или же иногда на стадии промежуточного продукта. Активирование карбоксильного компонента и следующее за ним образование пептидной связи, т. е. так называемая реакция конденсации, в идеальных условиях должны протекать с высокой скоростью без рацемизации, без побочных реакций и с высоким выходом при соединении эквимолярных количеств карбокси- и аминокомпонентов. К сожалению, в настоящее время еще неизвестно такого метода конденсации, который удовлетворял бы всем этим требованиям. Приходится выбирать из относительно большого набора методов подходящие варианты в соответствии со специфическими целями синтеза. Решение зависит в каждом случае от выбранной тактики синтеза, в соответствии с которой для каждого отрезка синтезируемой последовательности подбираются оптимальные методы конденсации. Набор методов, которые применяются для практического проведения синтеза пептидов, относительно мал по сравнению с примерно 130 описанными методами синтеза. На последней ступени пептидного синтеза происходит отщепление защитных групп. Поскольку синтез дипептида с полным удалением защитных групп проводится довольно редко, гораздо большее значение имеет селективное деблокирование, т. е. выборочное отщепление защитных групп N-концевой аминофункции или же С-концевой карбоксильной группы. Этот вопрос находится в тесной связи с общим планом синтеза.
Под стратегией понимают последовательность связывания аминокислотных компонентов в пептид, причем следует различать постепенное наращивание и фрагментную конденсацию. Получение полипептидов путем постепенного наращивания цепи трудноосуществимо при больших размерах целевой молекулы. В этих случаях большое значение приобретает разделение объекта синтеза на отдельные фрагменты с последующим соединением их в полипептид. Оптимальный выбор комбинации защитных групп и применение подходящего метода конденсации для каждого отрезка составляет предмет тактики пептидного синтеза.
Стратегическую модификацию постепенного наращивания пептидов или белков представляет разработанный в 1963 г. Меррифилдом пептидный синтез на полимерных носителях. Несмотря на сенсационный успех этого метода (синтез протекает в двухфазной системе и есть возможиость его автоматизации), возлагаемые на него большие ожидания до сих пор полностью не исполнились.
1.4 Экспериментальные методы создания пептидной связи
|
|
|
|
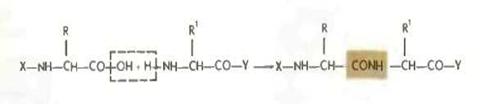
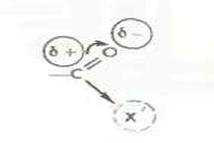
Образование пептидной связи в общем сводится к отщеплению элементов воды.
Для того чтобы сделать эту реакцию возможной и, более того, обеспечить ее высокую скорость и полноту, необходимо «активировать» карбоксильную группу. Такая активация должна сводиться к увеличению электрофильности карбонильного углерода[4].
Как легко видеть, важная роль в этом случае принадлежит группе X', которая в конечном счете определяет эффективность активации. Методы конденсации обычно и различаются природой группы X'.
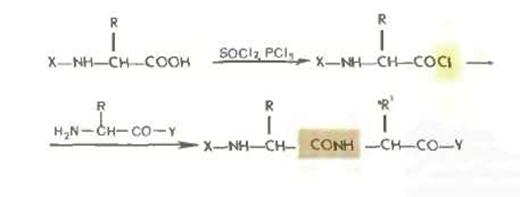
Хлорангидридный метод.
Хлорангидридный метод, упоминавшийся в историческом очерке, в настоящее время применяется редко, так как сопровождается рацемизацией и образованием побочных продуктов. Хлорангидриды получаются обычно обработкой производных аминокислот и пептидов хлористым тионилом или пятихлористым фосфором.
пептид молекула белок аминокислота
Азидный метод.
Метод Т. Курциуса находит широкое применение в синтезе пептидов.
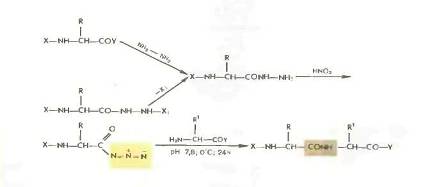
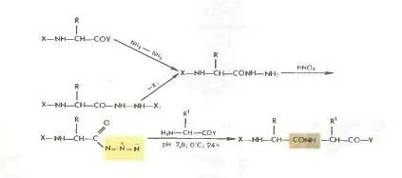
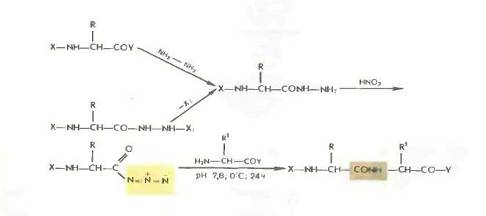
Гидразиды получаются либо прямым гндразинолизом эфиров защищенных аминокислот или пептидов, либо из защищенных гидразидов (—СО— NH — NH— Z—, - СО—NH и т. д.). Перевод в азиды осуществляется обработкой водным раствором нитрита натрия в кислой среде при —5 °С или действием изоамилнитрита или третбутил нитрита при —20 0 С в органическом растворителе (модификация Хонцля и Рудингера 1961). Азиды можно получать и непосредственно из НООС-производных с помощью днфенилфосфорилазида N3PO(OC2H5)z. применяемого в качестве конденсирующего агента (в частности, для получения циклопептидов).
Азидный метод не свободен от недостатков Основная побочная реакция — перегруппировка Курциуса:
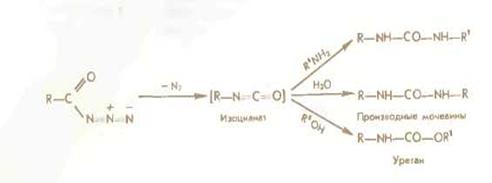
Направление и степень протекания этой перегруппировки определяется структурой азида и условиями реакции. Хотя при азидной конденсации рацемизация сведена к минимуму, ее нельзя не учитывать, особенно при блочном синтезе; в качестве основания рекомендуется использовать не триэтиламин-. а N-метилморфолин или N -этилдиизопропиламин.
Метод ангидридов.
Симметричные ангидриды ациламинокислот легко получаются обработкой последних дициклогексенкарбодиимидом или этоксиацетиленом например:
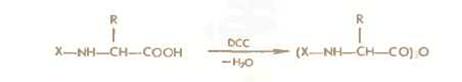
При реакции с аминокомпонентом образуются пептиды
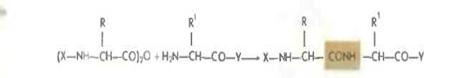
Метод в последнее время используется в твердофазном синтезе.
Более распространенным является применение смешанных ангидридов, в частности с производными угольной кислоты, получаемых с помощью изобутилхлоркарбоната (Р. Воган 1951).
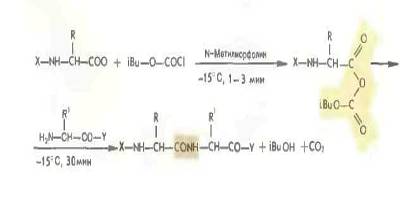
Быстрый ступенчатый метод синтеза пептидов с использованием избытка смешанных ангидридов носит название REMA-синтеза. К этой группе методов можно отнести и синтез пептидов с применением в качестве конденсирующего средства 1-этоксикарбо иил-2-этокси-1,2-дигидрохинолина.

Перспективны также смешанные ангидриды на основе производных ацилоксифосфония получае мые с помощью дифен ил фосфор ил хлорида (C2H5O)2POCl, тетра-этилпирофосфита (C2Р5O)2РООРО (OC2H5)2 или дифенилфос-фини хлорида (С Н ) РОС1 Основные побочные процессы при их использовании — это образование уретанов или оксазолонов и диспропорционирование
Метод активированных эфиров. Среди арильных активированных эфиров наиболее широки используются п- нитрофениловые (—ONр) (М. Боданскии 1956), 2,4 динитрофениловые, о-нит-рофениловые и о-нитротиофениловые, 2,4*5 трихлорфениловые (—ОТср), пентахлорфениловые (—ОРср). Особое значение приобрели недавно предложенные Л. Кишфалуди высокореакционноспособные пентафторфениловые эфиры (—OPfp). Ариловые эфиры этих типов получаются обычно из соответствующих фенолов с помощью карбодиимида.
В последние годы большое распространение получили активированные эфиры на основе производных гидроксиламина и прежде всего N- гидроксисукцинимидные эфиры, например:
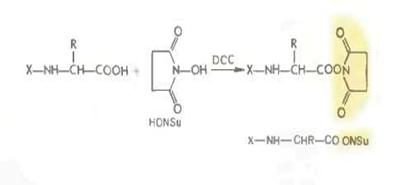
1.5 Первичная структура белка
В конце 50-х-начале 60-х годов было обнаружено, что последовательность аминокислот в белках детерминирована генетически. Последовательность нуклеотидов в ДНК-веществе наследственности определяет комплементарную последовательность нуклеотидов в РНК, а последняя в свою очередь определяет последовательность аминокислот в белке. Более того, синтез всех белков из соответствующих аминокислот имеет единый механизм.
Определение последовательности аминокислот в белках представляет интерес по четырем причинам. Во-первых, такие данные имеют большое значение для выяснения молекулярной основы биологической активности белка. Сведения о последовательности аминокислот приобретают особую ценность при их сопоставлении с другими химическими и физическими характеристиками. Во-вторых, изучение последовательности аминокислот в сочетании с детальным анализом пространственной структуры необходимо для выяснения тех принципов, на основе которых из полипептидных цепей формируются высокоспецифичные трехмерные формы. При этом последовательность аминокислот в белке служит связующим звеном между генетической информацией, заложенной в ДНК, и трехмерной структурой белка, лежащей в основе его биологической активности. В-третьих, изменения в последовательности аминокислот могут привести к нарушению нормальной функции белка, а следовательно, и к соответствующей болезни. В частности, такая тяжелая болезнь, как серповидно-клеточная анемия, нередко приводящая к легальному исходу, возникает в результате замены всею лишь одной-единственной аминокислоты в одном-единственном белке. Таким образом, определение последовательности аминокислот относится к новой области медицины -молекулярной патологии. В-четвертых, данные о последовательности аминокислот в белке могут многое рассказать о его эволюции. Дело в том, что в неродственных белках эти последовательности совершенно различны. Белки лишь в том случае имеют сходную последовательность аминокислот, если они происходят от общего белка-предшественника, Следовательно, изучение последовательностей аминокислот в белках позволяет проследить эволюцию на молекулярном уровне.
1.6 Экспериментальное определение первичной структуры белка
Рассмотрим, как можно определить последовательность аминокислот в коротком пептиде. Допустим, что пептид состоит из 6 аминокислотных остатков, расположенных в следующей последовательности:
Ala-Cly-Asp-Phe-Arg-Gly.
(Для обозначения аминокислот использованы общепринятые сокращения). Прежде всего необходимо определить аминокислотный состав пептида. Для этого его гидролизуют до составляющих аминокислот нагреванием до 110°С в течение 24 ч в 6 н. НС1. Далее аминокислоты полученного гидролиза разделяют методом ионообменной хроматографии на колонке с сульфонированным полистиролом. Фракционированные аминокислоты определяют по окраске, образующейся при нагревании с нингидрином: α-аминокислоты дают с нигидрином интенсивное синее окрашивание, а иминокислоты, например пролин, желтое[5].

Рис. 17 нингидрин и флуорескамин
Метод ионообменной хроматографии обладает высокой чувствительностью:
|
|

рН 3,25 0,2 м Цитрат натрия
рН 4,25 0(2 м Цитрат натрия
рН 5,28 0,35 м Цитрат натрия
Рис.18 Элюция из колонки
С его помощью можно определить даже один микрограмм аминокислоты, т.е. примерно столько, сколько содержится в одном отпечатке пальца. Количество аминокислоты пропорционально оптической плотности раствора после нагревания с нингидринйм. Если требуется определить еще меньшие количества аминокислоты- порядка нескольких нанограммов, то используют флуорескамин, который реагирует с α-аминогруппой, образуя сильно флуоресцирующее соединение. О природе аминокислоты судят по объему элюции, т.е. по объему буфера, использованному для вымывания данной аминокислоты с колонки (рис. ). Сравнение результатов хроматографии гидролизата со стандартной смесью аминокислот свидетельствует о том, что исследуемый пептид имеет следующий аминокислотный состав:
(Ala, Arg, Asp, Gly2, Phe).
Скобки показывают, что речь идет о составе, а не последовательности аминокислот в пептиде.
|
|
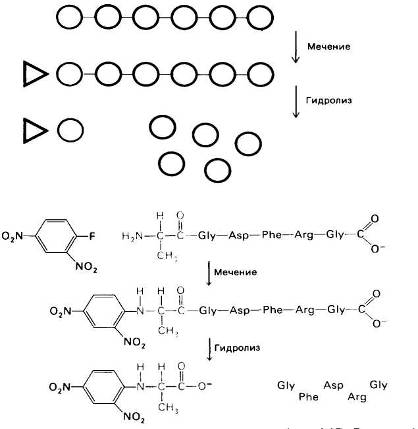
Рис.19 Определение N-концевого остатка пептида. Пептид метят фтординитробензолом (реактив Сэнгера) и затем гидролизуют. ДНФ-производное аминокислоты (в приведенном примере ДНФ-аланин) идентифицируют по хроматографиче-ским характеристикам.
Для определения в белке или пептиде концевого остатка, несущего аминогруппу, его метят с помощью соединения, образующего стабильную ковалентную связь с азотом аминогруппы (рис. 20).
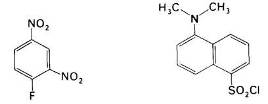
Рис. 20 фтординитробензол и дансилхлорид
Впервые для этой цели Сэнгер использовал фтординитробензол (ФДНБ), реагирующий с незаряженной a-NH2-группой с образованием динитрофенильного (ДНФ) производного пептида желтого цвета. Связь между ДНФ и концевой аминогруппой стабильна в условиях, используемых для гидролиза пептидных связей. Поэтому при гидролизе ДНФ-производного пептида Ala-Gly-Asp-Phe-Arg-Gly в 6 н. НСl высвобождается ДНФ-аминокислота, которую можно идентифицировать хроматографически как ДНФ-аланин.
Для идентификации N-концевых аминокислот в настоящее время часто используют дансилхлорид, который при взаимодействии с аминогруппой дает стабильное, интенсивно флуоресцирующее сульфамидное производное. Этот метод позволяет выявить N-концевую аминокислоту (после кислотного гидролиза пептидных
связей), присутствующую в таком незначительном количестве, как несколько нанограммов.
При всех достоинствах методов определения N-концевых аминокислотных остатков с помощью ДНФ или лансилхлорида их, к сожалению, нельзя использовать дважды применительно к одному и тому же пептиду, поскольку последний полностью распадается при кислотном гидролизе. Пьеру Эдману (P. Edman) удалось разработать метод маркирования N-концевого остатка и отщепления его от пептида без сопутствующего расщепления остальных пептидных связей. Деградация по Эдману (реакция Эдмана) состоит в ступенчатом (по одному) отщеплении аминокислотных остатков с аминоконца пептида, Фенилизотиоциа-нат реагирует с незаряженной концевой аминогруппой пептида с образованием фенилтиокарбамоильного производного
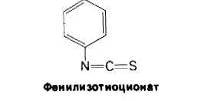
Далее в слабокислой среде происходит отщепление циклического производного N- концевой аминокислоты, а оставшийся неразрушенным пептид оказывается укороченным на один аминокислотный остаток, Указанное циклическое производное представляет собой фенилтиогидантоинаминокислоту (ФТГ-аминокислоту). Его идентифицируют методом хроматографии. Далее аминокислотный состав укороченного пептида (Arg, Asp, Gly2, Phe) сравнивают с исходным: (Ala, Arg, Asp, Gly2, Phe).
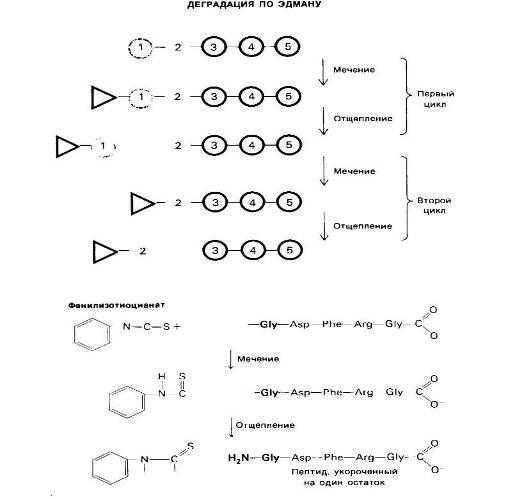 |
Рис 22. Деградация по Эдману.
От пептидной цепи отщепляют меченый N-концевой остаток аминокислоты (ФТГ-аланин на первой ступени деградации). Остаток пептидной цепи при этом не гидролизуется. На второй ступени деградации определяют следующий N-концевой аминокислотный остаток. Еще три ступени деградации но Эдману позволят установить всю последовательность аминокислот во взятом пептиде.
Оказывается, что различие состоит в одном остатке аланина. Следовательно, в исходном пептиде аланин занимает N-концевое положение. Деградацию по Эдману можно вновь повторить па укороченном пептиде. Исходя из аминокислотного состава после второй ступени деградации
(Arg, Asp, Gly, Phe)
Можно поийти к выводу, что вторым остатком с N-конца является глицин, это заключение подтверждают путем хроматографической идентификации ФТГ-глицина, полученного на второй ступени деградации пептида. Еще три ступени деградации по Эдману позволяют полностью раскрыть последовательность аминокислот во взятом пептиде.
Стратегию анализа последовательности аминокислот в белках можно определить как «разделяй и властвуй». Белок подвергают специфическому расщеплению па более короткие пептиды, последовательность аминокислот в которых определяют по Эдману. Специфическое расщепление можно производить химическими или ферментативным методами. Так, Б. Уиткоп (В. Witkop) и Э. Гросс (Е. Gross) обнаружили, что бромистый циан (CNBr) расщепляет полипептидную цепь только по пептидной связи, образованной карбоксильной группой остатка метионина, Если в белке содержится 10 метиониновых остатков, то после обработки бромистым цианом обычно получается 11 пептидов. Высокоспецифическое расщепление достигается также с помощью трипсина-протеолитического фермента поджелудочной железы.
Трипсин расщепляет полипептидные цепи по пептидной связи, образованной карбоксильной группой остатков аргинина и лизина. В результате белок, содержащий 9 остатков лизина и 7 остатков аргинина, после расщепления трипсином распадается на 17 пептидов. Каждый из этих пептидов, кроме пептида, расположенною на карбоксильном конце белка, будет кончаться аргинином или лизином.
Пептиды, полученные при специфическом химическом или ферментативном расщеплении белка, разделяют методами хроматографии. Далее последовательность аминокислот в каждом из пептидов определяют методом Эдмана. Таким образом, достигается этап, когда последовательность аминокислот в отдельных пептидах (фрагментах белка) известна, но остается невыясненной последовательность самих пептидов. Последнюю устанавливают с помощью так называемых перекрывающихся пептидов. При этом используют уже не трипсин, а какой-либо фермент, расщепляющий полипептидную цепь в других участках, например химотрипсин, который расщепляет пептидные связи главным образом по карбоксильным группам ароматических и других больших неполярных аминокислотных остатков
|
|
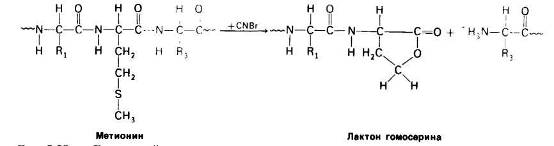
Рис. 23 Бромистый циан расщепляет полипептиды по карбоксильной группе метиониновых остатков.
Пептиды, образующиеся под действием химотрипсина, неизбежно перекрывают два или более триптических пептида, что используется для установления их последовательности. Таким путем полностью определяют последовательность аминокислот в белке.
Описанные методы применимы к белкам, состоящим из одной полипептидной цепи, не имеющей дисульфидных связей. В тех же случаях, когда в белке имеются дисульфидные связи или более одной полипептидной цепи, то необходимы дополнительные методические приемы, Например, если белок содержит две или более полипептидные цепи, соединенные нековалентными связями, то, воздействуя денатурирующими агентами, такими, как мочевина или гуанидингидрохлорид, вызывают диссоциацию цепей. Диссоциированные цепи разделяют и только после этого приступают к определению последовательности аминокислот в каждой из них. Если же полипептидные цепи соединены ковалентными дисульфидными связями, как это имеет место в инсулине, то их окисляют надмуравьиной кислотой; при этом дисульфидные связи разрываются и образуются остатки цистеиновой кислоты.
Анализ структуры белков удалось значительно ускорить путем создания секвенатора-специального прибора для автоматического определения последовательности аминокислот. При таком определении белок в виде тонкой пленки помещают во вращающийся цилиндрический сосуд, где он подвергается деградации по Эдману. Реактивы и растворители проходят над иммобилизованной белковой пленкой, а высвобождающиеся ФТГ-аминокислоты подвергаются жидкостной хроматографии при высоком давлении и таким образом идентифицируются. Один цикл деградации по Эдману занимает при этом менее двух часов. С помощью секвенатора можно определить аминокислотную последовательность полипептида или белка, содержащего до ста аминокислотных остатков.
ВЫВОДЫ
Данная курсовая работа обобщает материал о методах синтеза пептидов и о первичной структуре белков.
В работе приведены данные о структуре молекул белков, их строении, номенклатуре и методах синтеза; описаны как общие теоретические положения, так и экспериментальные методы, позволяющие для реальных систем реализовать на практике конкретные задачи.
Указаны трудности, связанные с проблемой синтеза пептидов. Проведен сравнительный анализ различных методов синтеза.
Описаны причины важности исследований в области первичной структуры. Приведена тактика определение первичной структуры белка, позволяющая определить аминокислотную последовательность, как небольших пептидов, так и молекул с числом остатков аминокислот, превышающим 100.
СПИСОК ИСПОЛЬЗОВАННОЙ ЛИТЕРАТУРЫ
1. Якубке Х.-Д., Ешкайт Х. Аминокислоты, пептиды, белки: Пер. с нем.- М.: Мир, 1985 г.-456 с.
2. Страйер Л. Биохимия: Пер. с англ.- М.: Мир, 1984.-Т.1-232 с.
3. Овчинников Ю.А. Биоорганическая химия.- М.: Просвещение, 1987.- 815с.
4. Тюкавкина Н.А., Бауков Ю.И. Биоорганическая химия: М.: Медицина, 1991.-528с.
5. А. Уайт, Ф. Хендлер Основы биохимии: М.: Мир Т.1, 1981.- 534 с.
