

 Курсовая работа: Загальна характеристика і особливості життєдіяльності бактерій родини Enterobacteriaceae
Курсовая работа: Загальна характеристика і особливості життєдіяльності бактерій родини Enterobacteriaceae
ГЦ-склад ДНК ентеробактерій варіює в широких межах – від 37 до 63%. За виключенням роду Proteus, представники якого суттєво відрізняються за нуклеотидним складом. ГЦ-склад ДНК споріднених організмів трьох видів - Salmonella, Escherichia і Shigella – розрізняються зовсім мало. Діапазон змінення
ГЦ-складу для „класичних” бактерій родини Enterobacteriaceae (від 37 до 59%) дуже близький до діапазону для представників родини з полярним джгутикуванням (від 39 до 63%) [10].
Бродіння
Зброджування сахаридів у ентеробактерій відбувається за шляхом Ембдена-Мейєргофа. Продукти бродіння у різних представників групи розрізняються і якісно, і кількісно. Однак у цих процесів є одна характерна біохімічна властивість, яка рідко зустрічається при інших типах бактеріального бродіння. Мова йде про особливий спосіб розщеплення однієї з проміжних сполук – піровиноградної кислоти (ПВК), що веде до утворення мурашиної кислоти:
СН3СОСООН + КоАSН→СН3СОSКоА+ НСООН.
Таким чином, мурашина кислота часто є основним кінцевим продуктом бродіння. Вона накопичується, хоча не завжди, оскільки деяки з цих бактерій володіють формнатгідрогенліазою, яка розщеплює мурашину кислоту до СО2 і Н2:
НСООН → СО2+Н2.
У таких організмів замість мурашиної кислоти як кінцевого продукту бродіння утворюються еквімолярні кількості Н2 і СО2.
Найбільш розповсюдженим типом перетворення сахаридів при бродінні у ентеробактерій є так зване бродіння змішаного типу, яке приводить до утворення молочної, оцтової, бурштинової і мурашиної кислот (або СО2 і Н2), а також етилового спирту. Такий тип бродіння властив представникам родів Salmonella, Escherichia, Shigella і Proteus. Відношення кількості різних кінцевих продуктів різниться як від штаму до штаму, так і для одного штаму при рості в різних умовах, наприклад при різних значеннях рН. Така різниця відображає ту обставину, що кінцеві продукти утворюються з піровиноградної кислоти трьома незалежними шляхами (рис. 3.1). Утворення газу в результаті зброджування сахаридів є дуже важливою ознакою для ентеробактерій – газоутворювальні бактерії роду Escherichia відрізняються за цим критерієм від патогенних видів групи Shigella і Salmonella typhi, які зброджують сахариди без виділення газу. В випадку простого бродіння змішаного типу газ може утворюватися тільки за рахунок розщеплення мурашиної кислоти; відповідно, утворення газу відображає наявність форміатгідрогенліази. Цей ферментний комплекс, звісно, не суттєвий для бродіння і може загубитися в результаті мутації без зміни здатності бактерії до бродіння. І дійсно, дослід показує, що в природі існують „неаерогенні”, тобто не утворюючі газоподібних продуктів штами такого типового продуценту їх, як Escherichia coli. Тому, хоча виділення газу являє собою корисну ознаку при класифікації ентеробактерій, даний критерій не є безпомилковим.
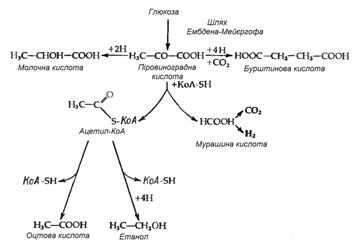
Рисунок 3.1 - Шляхи утворення із ПВК типових кінцевих продуктів кислотного бродіння змішаного типу (за [10])
Іншою ознакою, яка має велике значення для ідентифікації ентеробактерій, є здатність зброджувати дисахарид лактозу, що обумовлено наявністю у цих організмів β-галактозідази. Ефективність зброджування лактози залежить ще і від наявності специфічної пермеази галактозидів, яка сприяє проникненю лактози в клітину. Штами які мають β-галактозідазу, але позбавленні пермеази, не можуть поглинати лактозу зі швидкістю, яка достатня для інтенсивного бродіння, і зазвичай класифікуються як не здатні до зародження цього сахариду. Зброджування лактози властиво Escherichia, але відсутні у Salmonella, Shigella і Proteus. Деякі штами Shigella утворюють β-галактозідазу, але не можуть зброджувати лактозу із-за відсутності пермеази [10].
4. ГЕНЕТИКА БАКТЕРІЙ РОДИНИ ENTEROBACTERIACEAE
Відкриття явища переносу генів у ентеробактерій при кон’югації і трансдукції дозволило доволі детально дослідити генетичні родинні відносини між деякими її представниками. Утворення хромосомних гібридів між E. coli і бактеріями родів Salmonella і Shigella каже про дуже високу ступінь генетичної гомології цих бактерій, що підтверджується і порівнянням хромосомних карт двох найбільш вивчених в цьому відношенні видів − Salmonella typhimurium і Escherichia coli. Як видно на рис. 4.1 і 4.2, де представленні в спрощеному вигляді хромосомні карти цих двох організмів, багато маркерів розташовуються на обох хромосомах в однакових локусах.
Про близьку генетичну спорідненість груп Escherichia, Salmonella і Shigella свідчить також висока ступень ДНК-ДНК-гібридизації in vitro. Утворення ж хромосомних гібридів між бактеріями цієї підгрупи і представниками інших родів ентеробактерій (Proteus, Enterobacter) відбувається дуже рідко. Крім того, випробування по гібридизації ДНК-ДНК вказують, що ступень генетичної гомології між представниками підгрупи Escherichia− Salmonella− Shigella та іншими групами ентеробактерій доволі низька. Ентеробактерії, що належать до різних родів, можуть отримувати плазміни від донорних штамів E. coli при кон’югації і потім зберігати їх як позахромосомні елементи (ефективність кон’югації варіює в широких межах; таблиця 4.1). Таким чином серед бактерій кишкової групи можуть розповсюджуватися F-фактори, які містять додаткові гени (наприклад F-lac), і R-фактори, які визначають стійкість до різноманітних лікарських засобів [10].
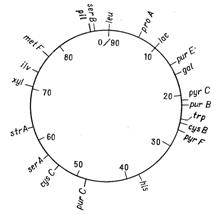
Рисунок 4.1 - Генетична карта E. coli К12 в спрощеному вигляді (за [10])
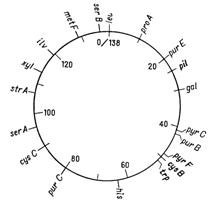
Рисунок 4.2 - Генетична карта S. typhimurium в спрощеному вигляді (за [10])
5. ЕКОЛОГІЯ БАКТЕРІЙ РОДИНИ ENTEROBACTERIACEAE
Середовищем проживання для більшості ентеробактерій є кишковий тракт хребетних тварин і людини.
В організмі людини багато ентеробактерій містяться в складі мікробних біоценозів тонкого і товстого кишечника. Патогенні види зустрічаються тільки у хворих і бактеріоносіїв.
З випорожненнями людей і тварин ентеробактерії потрапляють в навколишнє середовище. В ній вони можуть зберігатися на протязі різних строків в залежності від виду і умов. Деякі з них (E. coli) використовуються в санітарній мікробіології в якості показників фекального забруднення навколишнього середовища [3].
Кишкова паличка наприклад може зберігатися в воді і ґрунті декілька місяців [11].
Широке розповсюдження сальмонел в природі і різноманіття шляхів, якими вони проникають в організм людини, пояснюється їх генетичною пластичністю. Сальмонели легко знаходять екологічні ніші і адаптуються до найрізноманітніших умов [12].
Клебсієли – широко розповсюджені бактерії. Вони порівняно стійкі до факторів навколишнього середовища завдяки наявності капсули і можуть на протязі тривалого часу зберігатися в ґрунті, воді, приміщеннях. Разом з тим вони зустрічаються в складі мікробних біоценозів в організмі людини і тварин [3].
Протеї P. vulgaris і P. mirabilis є мешканцями кишечника багатьох тварин, виявляються також в стічних водах і ґрунті. Всі види протеїв можуть виділятися з хворих людей [3].
6. АНТИГЕНИ БАКТЕРІЙ РОДИНИ ENTEROBACTERIACEAE
Антигенна будова служить одним із суттєвих критеріїв, на яких основана класифікація, а також ідентифікація ентеробактерії. Розрізняють три основних типи антигенів: 1) О-соматичний антиген; 2) Н-джгутиковий антиген; 3)К-антигени. О-антиген є складовою частиною ліпополісахариду (ЛПС) зовнішнього шару клітинної стінки. Специфічність О-антигену визначається детермінантними сахаридами (гексозами і аміносахаридами), ковалентно зв’язаними з базисною частиною ЛПС. Н-антиген локалізується в джгутиках клітини. Він складається з білка флагеліну. Капсульні антигени К-антигени, так як і О-антигени, містяться в ЛПС клітинної стінки, але в більш поверхневому шарі. Вони маскують О-антигени.
В зв’язку з цим для визначення О-антигену у бактерій, які володіють К-антигеном, необхідно зруйнувати останній шляхом кип’ятіння або автоклавування культури. К-антигени за хімічними властивостями відносяться до кислих полісахаридів. До К-антигенів відносяться К-антигени E. coli, М-антигени Salmonella schottmuelleri, Vi-антиген збудника черевного тифу, який виявляється також у S. parathyphi C та деяких штамів E. coli. Всі ці антигени характеризуються імуннохімічною специфічністю, що дозволяє диференціювати роди і види, а також виділяти серед них серогрупи і серологічні варіанти (серовари). Антигенну будову ентеробактерій вивчають в реакціях аглютинації, непрямої (пасивної) гемаглютинації, преципітації, зокрема при імунно-електрофорезі та інших імунологічних реакціях з відповідними діагностичними сироватками.
Крім антигенів які перелічені, у ентеробактерії виявленні загальні антигени. Один з них (антиген Куніна) зв’язаний з ЛПС, інший – з білками клітинної стінки. Антиген Куніна подібне до антигену, який виявлений в товстому кишечнику людей, які страждають виразковим колітом [3].
Ешерихії Кишкова паличка має складну антигену структуру. Вона містить соматичний, або О-антиген, поверхневий (капсульний, оболонковий) К-антиген і джгутиковий Н-антиген. О-антигени у бактерій триби має схожу хімічну будову і зв’язані з ЛПС клітинної стінки. Н-антигени є тільки у джгутикових форм бактерій і складаються з білка флагеліну.
О-антигени є основними антигенами які визначають серологічну групу ешеріхій. В теперішній час описано близько 170 О-серогруп E. coli. Більшість ешеріхій, що належать до різних О-серогруп, зв’язані одна з одною перехресними антигенними зв’язками. Близько 100 серогруп мають антигенні зв’язки з шигелами, сальмонелами та іншими ентеробактеріям.
К-антигени представленні трьома антигенами, які позначаються літерами A,B і L. Вони відрізняються один від одного чутливістю до нагрівання і хімічним речовинам. К-антигени мають здатність маскувати О- антигени, які можна виявити тільки після руйнування перших кип’ятінням культури. Ешерихії містять близько 97 різних К-антигенів, переважно В-типу.
Н-антигени на відміну від О- і К-антигенів є типоспецифічними. У ешеріхій описано близько 50 різних Н-антигенів.
Антигени ешеріхій позначаються антигенними формулами, які вказують на серогруп, наприклад E. coli О126:К6О(В6), або серовар − E. coli О126:К6О(В6):Н2 [3,13].
Сальмонели
Сальмонели, так як і ешерихії, мають складну антигенну структуру. Вони містять О-антигени (соматичні) та Н-антигени (джгутикові). Деякі сальмонели мають К-антиген. За своєю хімічною структурою О-антигени сальмонел аналогічні тим же антигенам ешерихій. Вони відрізняються один від одного тільки структурою кінцевих ланок полісахаридного компоненту ЛПС, який визначає імуннохімічну специфічність грамнегативних бактерій. Н-антигени можуть існувати в двох різних фазах: специфічній 1-й фазі і менш специфічній, або груповій, 2-й фазі. Систематична розробка антигенної будови різних сальмонел була проведена Ф. Кауфманом, який в 1934 році запропонував серологічну класифікацію цих бактерій. Всі відомі на той час сальмонели (близько 700) він розділив на 44 серологічних типи в реакціях аглютинації з діагностичними моно рецепторними сироватками.
Ця схема в подальшому неодноразово розширювалася Ф. Кауфманом та П. Уайтом і зараз включає близько 2000 сероварів сальмонел, які входять в склад 4 підродів.
До підроду I − S. kauffmani − відноситься більша частина патогенних для людини сальмонел серологічних груп A, B, C, D, E.
Аналіз антигенної будови є обов’язковим елементом мікробіологічної діагностики сальмонельозів. В основу схеми Ф. Кауфмана і П. Уайта закладена общин О-антигенів сальмонел, які об’єднанні в серологічні групи, які позначаються літерами латинського алфавіту. Диференціація сальмонел всередині групи проводиться на основі особливостей їх Н-антигенів. Фаза 1 позначається прописними літерами латинського алфавіту: a, b, c тощо, фаза 2 − арабськими числами і, рідше, латинськими літерами. Кожному сировару привласнюють одна з видових назв з відповідною антигенною формулою, наприклад: S. enteritidis 1, 9, 12 [3].
Сальмонели − збудники черевного тифу і паратифів А і В (S. typhi, S. parathyphi A, S. schottmuelleri)
Сальмонели черевного тифу і паратифу разом з О- і Н-антигенами можуть містити і Vi-антиген. Він відноситься до К-антигенів і являє собою полімер N-ацетилгалактозаміноуронової кислоти. Наявність у сальмонел Vi-антигену, так як і ешерихій К-антигену, перешкоджає аглютинації бактерій О-сироватками. Vi-антиген міститься в вірулентних штамах сальмонел і є специфічним рецептором для деяких фагів, які називаються Vi-фагами. Це дало змогу розробити методику визначення фаговарів черевнотифозних і паратифозних сальмонел і розподілити їх по певним групам. Визначення фаговару згаданих сальмонел, так як і серовару, проводиться для епідеміологічного аналізу черевного тифу і паратифів з метою встановлення джерела інфекції, шляхів і засобів її розповсюдження [3].
Шигели так як і ешерихії і сальмонели мають складну антигенну структуру. В складі їх клітинних стінок є О-, а у деяких видів (шигели Флекснера) і К-антигени. За хімічною структурою вони аналогічні антигенам ешеріхій. Відмінності складаються головним чином в структурі кінцевих ланок полісахаридного компоненту ЛПС, які обумовлюють імуннохімічну специфічність, що дає змогу диференціювати їх від інших ентеробактерій і між собою. Крім того, шигели мають перехресні антигенні зв’язки з багатьма серогрупами ентеропатогенних ешерихій, які викликають головним чином дизентерієподібні захворювання, і з іншими бактеріями [3].
Клебсієли
Клебсієли містять О- і К-антигени. Всього відомо 11 О-антигенів і 80 К-антигенів. Останні зв’язані з капсулами. Серологічна класифікація клебсієл основана на їх антигенних відмінностях. У штамів однієї і тієї ж О-серогрупи можуть бути різні К-антигени і навпаки. Деякі О- і К-антигени клебсієл споріднені О-антигенам ешерихій і сальмонел. Найбільша кількість О- і К-антигенів знайдено у K. pneumoniae. За своїм хімічним складом ці антигени принципово не відрізняються від подібних антигенів інших ентеробактерій [3].
Протеї
Протеї володіють принаймні двома антигенами. Один з них представлений джгутиковим Н-антигеном, а другий – О-антиген − є ЛПС клітинної стінки. У трьох видів – P. morganii, P. rettgeri, P. inconstans ідентифіковано декілька десятків сероварів.
Деякі
серовари протея (ОХ-штами) мають антигени, які перехресно реагують з антигенами
рикетсій. Раніше ця властивість використовувалась при серодіагностиці сипного тифу
– реакція Вейля-Фелікса. В якості антигенів для цієї реакції слугували культури
Proteus OX![]() [3].
[3].
7. ПАТОГЕНІСТЬ І ТОКСИНОУТВОРЕННЯ БАКТЕРІЙ РОДИНИ ENTEROBACTERIACEAE
Патогенна дія ентеробактерій пов’язана з ліпополісахаридами клітинної стінки.
Вірулентність ентеробактерій визначається їх адгезивною здатністю, яка обумовлена позитивним хемотаксисом між поверхневими структурами мікробу і рецепторами епітеліальних клітин. Крім того, адгезивність пояснюється наявністю у багатьох ентеробактерій ворсинок, а також структурою ЛПС. Після „прилипання” до епітеліальних клітин ешерихії, які викликають дизентерієподібні колі-інфекції, і шигели проникають всередину цих клітин і там розмножуються: сальмонели розмножуються в макрофагах лімфоїдної тканини тонкого кишечника (в пейерових бляшках) і внутрішніх органах, а ешерихії – на поверхні клітин. Потім бактерії поступають у просвіт кишечника.
Токсигенність ентеробактерій обумовлена ендотоксином і екзотоксинами (ентеротоксинами і цитотоксинами). Перший являє собою ЛПС клітинної стінки. Він вивільняється тільки після руйнування бактеріальних клітин. Ентеротоксигенні ешерихії продукують два типи ентеротоксину білкової природи, які розрізняються чутливістю до температури. Їх утворення контролюється Ent-плазмідою. Термолабільний ентеротоксин, який утворюється ешерихіями і шигелами, за своїми властивостями схожий на холероген, який продукується холерним вібріоном. Багато ешерихій, які викликають колі-ентерити і дизентерієподібні захворювання, а також шигели Флекснера, Зонне, сальмонели тощо, здатні утворювати ентеротоксини [3].
До дійсного часу систематизовано біля ста патогенних серотипів кишкової палички, що викликають захворювання в людини, тварин, у тому числі і птахів.
З представників групи кишкової палички найбільш патогенної вважають підгрупу A. aerogenes (І.С.Загаєвський). Ці бактерії часто викликають колібактеріоз у телят і дітей, важкі мастити в корів, гостре запалення легких і сечостатевих шляхів у людини і тварин. Крім захворювання, деякі види бактерій кишкової палички викликають псування молока і молочних продуктів [14].
